По-видимому, большое значение в процессах регуляции клеточного деления имеет группа белков. программируемых так называемыми онкогенами. Измененные (мутантные) формы этих генов обнаруживаются в опухолевых клетках и входят в ряде случаев в виде соответствующих РНК-копий в состав онкогенных (т.е. вызывающих опухоли) ретровирусов. Первым открытым онкогеном был ген sr. входящий в состав вируса саркомы Рауса. Программируемый им белок, продукт гена sr. оказался протеинкиназой, которая в отличие от протеинкиназ класса А и протеинкиназы С катализировала фосфорилирование определенного спектра клеточных белков по остаткам тирозина, а не по остаткам серина и треонина, Дальнейшие исследования показали, что такая активность присуща некоторым рецепторам факторов роста. в частности рецептору эпидермального фактора роста. Ген erd, программирующий аналог этого рецептора, был обнаружен в составе онкогенного вируса птичьего миелобластоза, В настоящее время открыто несколько десятков онкогенов. В большинстве изученных случаев продукты этих онкогенов в здоровых клетках являются участниками передачи митогенных (т. е. управляющих, митозами) сигналов. В ряде опухолей, в том числе человеческих, найдены онкогены, программирующие аналоги белка G,воспринимающего сигна-, лы от комплексов эффектор - рецептор (в частности, онкогены Н—ras и К—ras) онкогены, программирующие синтез аналогов самих факторов роста. например онкоген sis, входящий в состав вируса саркомы обезьян, продукт которого является аналогом фактора роста. выделяемого тромбоцитами (клетками крови. участвующими в процессе свертывания ) онкогены, продуктами которых являются аналоги ядерных белков, по-видимому, участвующих на заключительных этапах каскада превращений. возникающего в ответ на митогенный сигнал (онкогены туе, fos и др.). [c.428]
Молекулярные веса вирусных ДНК колеблются в пределах от 1,3 10 до (120—130) 10 (для фага Т 2), а молекулярные веса РНК для фага — от 1,3-10 до 2-10. однако имеются некоторые отклонения до (9—10)10 (реовирусы, вирус саркомы Рауса и др.)- [c.467]
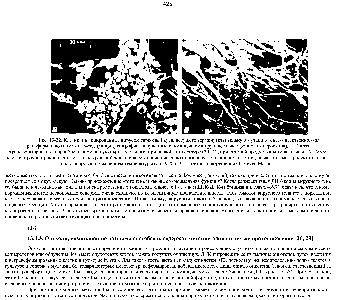
Рис. 13-32. Клетки, инфицированные вирусом саркомы Рауса. несущего термочувствительную мутацию гена, ответственного за трансформацию (онкогена -sr ) (микрофотографии, полученные с помощью сканирующего электронного микроскопа ). А Клетки трансформированы и приобрели аномальную округлую форму при низкой гемпературе (34 °С), при которой продукт онкогена активен. Б. Те же клетки, прочно прикрепленные к культуральной чашке и восстановившие свою нормальную уплощенную форму. после того как продукт онкогена
РНК опухолеродных вирусов. таких, как вирус саркомы Рауса. содержат РНК с молекулярным весом приблизительно 10-10. которая ведет себя как единый компонент в условиях различных ионных сил, включая даже те условия, при которых РНК вируса гриппа диссоциирует. Тем не менее при нагревании или в присутствии диметилсульфоксида — растворителя, нарушающего взаимодействие между основаниями. происходит резкое изменение седиментационной картины появляется главный компонент с молекулярным весом около 3 -10 и более мелкие фрагменты РНК [99]. [c.114]
Некоторые виды опухолей вызываются вирусами. Выделенные из них вирусы инфицируют нормальные клетки и, внося в них свою РНК или. ДНК, трансформируют эти клетки в опухолевые. Первым из таких опухолеродных (онкогенных) вирусов был вирус саркомы Рауса. вызывающий соединительнотканные опухоли (саркомы) у птиц. Он стал важным объектом исследований. так же как и ряд других онкогенных вирусов. открытых позже. [c.426]
Изучение вируса саркомы Рауса. как обычно, лишь тогда увенчалось успехами, когда был разработан надежный метод его обнаружения, а именно упомянутое выше образование доступных для подсчета гроздьев, или фокусов. трансформированных клеток в культуре монослоя фибробластов цыпленка [498]. В отличие от того, что [c.273]
Тот факт, что опухоли, вызываемые вирусом саркомы Рауса. сильно различаются по содержанию в них вирусов, может быть обусловлен одним из двух различных механизмов. С одной стороны, опухоли, индуцируемые большими дозами вируса у взрослых птиц, постепенно теряют большую часть своих инфекционных вирусов благодаря иммунологическому ответу хозяина. Эту реакцию можно [c.275]
Недавно были найдены условия, в которых определенные штаммы вируса саркомы Рауса индуцируют у млекопитающих развитие опухолей [5]. И в этом случае чувствительными к инфекции оказались новорожденные животные, и особенно хомячки. Эти опухоли не продуцируют инфекционных частиц вируса саркомы Рауса [177, 178], пока их не переносят обратно на цыплят. [c.276]
Когда ретровирус трансформирует нормальную клетку в опухолевую, аномальное поведение часто бывает обусловлено геном, который ( привнесен вирусом, но для выживания и репродукции самого вируса фактически не нужен. Впервые это выяснилось, когда были открыты мутанты вируса саркомы Рауса. способные нормально размножаться, но не трансформирующие клетку. Оказалось, что некоторые из этих нетрансформирующих мутантов не имеют гена (или части гена ), кодирующего белок с мол. массой 60000. В результате других мутаций этого гена трансформирующее действие вируса может становиться термочувствительным зараженные клетки проявляют трансформированный фенотип при 34°С, но после повышения температуры до 39°С они быстро (через несколько часов) возвращаются к нормальному фенотипу (рис. 13-32). По-видимому, этот специфический ген в онкогенном вирусе ответствен за клеточную трансформацию (и этим привлекает наше внимание), но является ненужным балластом с точки зрения репродукции самого вируса. [c.427]
Одним из наиболее интенсивно изучаемых протоонкогенов является -sr. соответствующий онкогену л -sr вируса саркомы Рауса. Он принадлежит к небольшому семейству гомологичных протоонкогенов и кодирует белок sr - тирозин-специфическую протеинкиназу с мол. массой 60000 (поэтому иначе ее называют РбО ), которая содержит ковалентно связанную жирную кислоту. прикрепляющую ее к внутренней стороне плазматической мембраны (разд. 8.2.3). В своей онкогенной форме эта киназа сверхактивна, и для того, чтобы она могла вызывать трансформацию клетки. необходимо ее прикрепление к мембране (рис. 13-35). Эксперименты с антителами показывают, что [c.431]
А, транскрипция которой находилась под контролем промотора вируса саркомы Рауса или ци-томегаловируса. Хотя уровень экспрессии гена нуклеопротеина был настолько низок, что не поддавался регистрации, через 2 нед после иммунизации в крови мышей обнаруживались антитела к нему. Выживаемость иммунизированных мышей оказалась значительно выше, чем мышей из контрольной группы (рис. 11.5). Более того, они были нечувствительны и к другому штамму вируса гриппа. Такая перекрестная защита не вырабатывается при введении традиционных противогриппозных вакцин. полученных на основе поверхностных антигенов вируса. и поэтому каждая вакцина специфична лишь к одному штамму вируса. Более того, традиционные вакцины сохраняют свою эффективность только до тех пор, пока остаются неизмененными поверхностные антигены. К сожалению, для генов поверхностных антигенов характерна высокая частота мутаций, что приводит к появлению существенно различающихся штаммов вируса. Кбровые же белки, такие как нуклепротеин, относительно стабильны и активируют иммунную систему по другому механизму. чем поверхностные антигены. [c.233]
Рис. 11.5. Выживание мышей, иммунизированных вирусной ДНК. Тестируемых мышей иммунизировали Е. со//-плазмидой, несушей кДНК нуклеопротеина вируса гриппа А под контролем промотора вируса саркомы Рауса. Контрольным мышам вводили только плазмидную ДНК. По оси абсцисс отложено время, прошедшее после контакта животных с вирусом гриппа.
Способность фенолов задерживать и инициировать радикальные реакции объясняет многообразие их свойств. При воздействии ПГ была установлена потеря бластомогенных свойств вируса саркомы Рауса [69], Обнаружено подавление пропилгаллатом вируса ядер-ного полиэдроза дубового шелкопряда [70]. При этом Кок [71 показала, что под действием ПГ в тканях куг олок шелкопряда уменьшается количество нуклеиновых кислот. Известно, что фенольные ооединения подавляют развитие растительных раков. Так, ПГ подавляет развитие рака картофеля, вызванного Ba terium iumifd iens [72], Подобный эффект был получен на опухолях подсолнечника и томатов, вызванных теми же бактериями [73], [c.328]
Б. Миграция в 0,1%-пом растворе додецилсульфата натрия (pH 7,2). В обоих случаях Пелок ВТМ добавлен в качестве маркера, поглощающего ультрафиолет. Белки вируса саркомы Рауса обнаруживаются поих радиоактивности [100]. [c.85]
Недавно было показано, что вирус саркомы Рауса также содержит несколько компонентов РНК с молекулярным весом до 3-10. а выделенная ранее РНК с константой седиментации 708 представляет собой агрегат этих компонентов. В этом случае комплекс не диссоциирует под действием ЭДТА, однако диссоциация происходит при нагревании и обработке РНК диметилсульфоксидом. Объясняется это, по-видимому, тем, что отдельные компоненты РНК этого вируса и других сходных с ним опухолевых вирусов удерживаются вместе в результате возникновения водородных связей между определенными комплементарными друг другу участками этих молекул (см. гл. VI, разд. Г) [99]. РНК вируса саркомы Рауса. выделенная в виде комплекса или в диссоциированном состоянии, также не обладает инфекционностью. [c.163]
Совсем по-иному складываются отношения между дефектным вирусом. например вирусом саркомы Рауса. и вирусом-помощником. В этом случае неспособность вируса кодировать синтез какого-либо (функционального) белка компенсируется образованием этого белка вирусом -помощником (см. гл. ХИ, разд. В) [396]. Также отличается по своему характеру роль вируса -помощника в процессе инфицирования клеток Е. oli с помощью ДНК фага "К. В обоих этих случаях нуклеиновая кислота сохраняет способность синтезировать свою репликазу в отличие от нуклеиновой кислоты вируса -сателлита вируса некроза табака. [c.169]
Случаи, когда у изолированных нуклеиновых кислот не удается обнаружить инфекционности, можно объяснить двумя основными причинами. Во-первых, вполне возможно, что молекулы РНК и ДНК с молекулярным весом от 6-10 до 200-10. примерами которых могут служить PFIK вируса ньюкаслской болезни (молекулярный вес 6-10 ), ДНК аденовирусов (молекулярный вес 22-10 ) и Д11К вируса осповакцины (180-10 ) не способны проникнуть в клетку в интактном состоянии без помощи защитного капсида или наружной оболочки вируса. Во-вторых, как теперь известно, РНК некоторых других вирусов, содержащаяся в них в меньшем количестве, состоит из нескольких молекул. В этом случае вероятность проникновения в клетку-хозяина полного набора генетических компонентов. лишенных преимуществ специфической упаковки, сводится, должно быть. к минимуму. Возможно, что этим и объясняется отсутствие инфекционности у РНК вируса гриппа (вероятно, 6 компонентов с общим молекулярньш весом 3-10 ), у РНК вируса саркомы Рауса (3 компонента, каждый с молекулярным весом около 3-10 ) и у РНК реовируса (10 двухцепочечных компонентов с общим молекулярным весом 14-10 ). [c.173]
В 1970 году было точно доказано, что при включешги вируса саркомы Рауса в геном клетки-хозяина происходит синтез ДНК на РНК-матрнцо. (Т е m i п И. М. et al. Nature, 1970, 227).— Прим. ред. [c.266]
Вирус саркомы Рауса (вирусы Брайана, Шмпдта-Руппина и др.) Плотные опухоли (птицы) [395] [c.274]
По-видимому, все клетки, инфрщированные вирусом саркомы Рауса. трансформируются и становятся злокачественными [400]. Высказывалось предположение, что эта высокая степень онкогенности, не свойственная другим вирусам. связана с онисанными выше проявлениями дефектности у этого вируса. Однако теперь это кажется маловероятным, поскольку штаммы вируса саркомы Рауса. не обладающие псевдодефектными свойствами, также оказываются хорошими трансформантами то же самое относится к высокопродуктивному штамму Брайана, если он атакует чувствительные к нему клетки. [c.275]
Некоторые данные о репликации вируса саркомы Рауса служат довольно ясным намеком на механизм онкогенеза, индуцированного РНК-содержащими вирусами. Оказалось, что уже на ранних стадиях инфекции вирусом саркомы Рауса должен происходить синтез ДНК, и при этом необходимо, чтобы ДНК функционировала на протяжении всего процесса репликации вируса [24] — обстоятельство, прямо противоположное тому, что наблюдается с репликацией всех неонкогенных РНК-содержащих вирусов. Высказывалось предположение, что РНК — ДНК-гибрид может представлять собой эквивалент репликативной формы (РФ) этих онкогенных вирусов [24]. Существует также предположение об образовании двух -ценочечных ДНК-провирусов, включенных в геном хозяина [497] Опыты по гибридизации ДНК хозяина и РНК вируса саркомы Рауса или РНК какого-нибудь другого вируса лейкоза показали, что небольшие сегменты этих РНК, особенно богатые аденином, комплементарны к ДНК-хозяина 1180, 181]. [c.276]
В чем основное различие между высоким титром штамма Брайана и штаммом Шмидта — Рупина вируса саркомы Рауса [c.287]
Ретровирусы содержат молекулы одноцепочечной РНК, при этом каждая вирусная частица имеет две копии РНК-генома, т. е. вирусы этого типа являются единственной известной разновидностью диплоидных вирусов. Впервые они были обнаружены по способности вызывать образование опухолей у животных. Первый вирус этого типа был описан в 1911 г. Пейтоном Раусом, обнаружившим инфекционную саркому у кур. Вызывающий это заболевание вирус. названный вирусом саркомы Рауса (ВСР), послужил моделью для изучения не только опухолеродных РНК-вирусов, но и других ретровирусов, не приводящих к возникновению опухолей. ВСР может инфицировать культивируемые in vitro куриные эмбриональные клетки и превращать их в клетки с измененными свойствами. характерными для опухолевых клеток. Изме- [c.49]
Ретровирусы, вызывающие раковую трансформацию, это РНК-со-держащие вирусы. С помощью фермента ревертазы они способны синтезировать ДНК-копии в ходе так называемой обратной транскрипции. Эти ДНК-копии способны встроиться в геном клетки. Интегрированная копия называется провирусом. Некоторые ретровирусы, известные как активно трансформирующие вирусы. высоко онкогенны. Они вызывают неопластические заболевания у зараженных ими животных. В культуре клеток эти вирусы вызывают трансформацию клеток, протекающую с высокой эффективностью. Около 20 таких вирусов было выделено из крыс, мыщей, обезьян, кощек, цыплят и индюков (например, вирусы саркомы Харвея и саркомы Малони выделены из крыс и мышей соответственно). Кроме генетической информации. необходимой для своей собственной репликации, эти вирусы несут специфические гены. называемые онкогенами, ответственными за их способность вызывать раковую трансформацию. Сейчас известно около 15 генов one, включая ген sr вируса саркомы Рауса. поражающего кур, ген mos вируса саркомы мышей и ген ras вируса саркомы крыс. [c.322]
Дополнительным подтверждением гомологии между онкогенами и последовательностями нормальных клеток служит и тот факт, что они, как правило, кодируют аналогичные белки. Например, онкоген вируса саркомы Рауса кодирует тирозин-специфичную киназу. обозначаемую ррбО. Этот белок представляет собой фосфопротеин молекулярной массы 60000. Такой же белок выделяется и из нормальных клеток цыпленка, хотя в этих клеттсах его количество в 100 раз меньше, чем в раковых. Использование клонированных фрагментов онкогенов в экспериментах по ДНК-гибридизации выявило гомологию между геном газ вируса саркомы Харви и геном из клеток карциномы мочевого [c.323]
Особым образом ведут себя опухолевые РНК-вирусы проникновение их в пермиссивиую клетку часто ведет одновременно и к нелетальному для клетки высвобождению дочерних вирусных частиц (отпочковывающихся от клеточной поверхности ), и к стойкому генетическому изменению в инфицированной клетке. которое превращает эту клетку в раковую. Как может заражение вирусом вызвать стойкое генетическое изменение. было непонятно до тех пор. пока не был открыт фермент обратная транскриптаза с помошью которого пепи инфицирующей РНК этих вирусов транскрибируются в комплементарные им цепи ДНК. Опухолевые РНК-вирусы, к которым относится первый хорошо изученный опухолевый вирус. а именно вирус саркомы Рауса. являются представителями крупного класса вирусов. так называемых ретровирусов. Название это отражает тот факт, что часть их жизненного цикла составляет процесс. обратный нормальной транскрипции, т. е. транскрипции ДНК в РНК. К ретровирусам относгггся и вирус СПИДа (спонтанно приобретенного иммунодефицита). [c.320]
Рис. 13-35. Эта электронная микpoфoтoqзaфия показывает, что протеинкиназа, кодируемая онкогеном -sr вируса саркомы Рауса. прикреплена к внутренней поверхности плазматической мембраны как полагают, белок sr. образующийся под действием белка -sr. по-видимому, находится там же, но его труднее обнаружить, так как он обычно присутствует в очень малых количествах Локализация белка sr была определена на этом препарате по его реакции со специфическими антителами. к которым были присоединены электроноплотные частицы

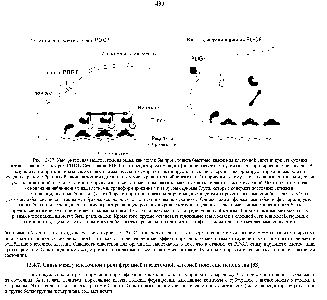
Рис. 13-37. Умозрительная модель, поясняющая, как могли бы происходить быстрые изменения клеточной адгезии при стимуляции клеток к делению фактором PDGF. Связывание PDGF с его рецептором приводит (пока не известным путем) к фосфорилированию белка -sr. В результате эта протеинкиназа, связанная с плазматической мембраной, активируется и в свою очередь фосфорилирует тирозиновые остатки соседних трансмембранных белков клеточной адгезии. в том числе рецептор фибронектина. Это приводит к тому, что фокальные контакты и другие участки клеточной адгезии частично разрушаются и связанные с ними актиновые филаменты теряют связь с мембраной. Частично эта модель основана на наблюдениях над клетками, трансформированными вирусом саркомы Рауса. которые содержат постоянно активный
И ДПК-, и РПК-содержащие вирусы (в частности, ретровирусы) могут участвовать в трансформации нормальной клетки в опухолевую. Это можно экспериментально продемонстрировать как на лабораторных животных (у которых некоторые вирусы способны вызывать рак), так и в культуре клеток, где те же вирусы изменяют поведение инфицированных клеток. Эти клетки приобретают способность к делению в условиях, при которых нормальные клетки делиться не могут (см. разд. 13.4.1, где перечислены свойства неопластически трансформированных клеток в культуре). Два интенсивно изучаемых примера таких вирусов - это 8У40 (ДПК-содержащий вирус, выделенный из клеток обезьяны) и вирус саркомы Рауса - куриный ретровирус. Сложнее обстоит дело с ролью вирусов в развитии рака у человека среди множества причин. приводящих нрактически ко всем известным видам раковых заболеваний человека, вирусы не фигурируют. Возможно, от многих вирус-индуцированных опухолей нас защищает иммунная система. разрушающая инфицированные вирусами клетки, которые могли бы стать источником опухолей. Тем не менее сейчас имеются веские доказательства того, что причиной возникновения некоторых типов рака человека являются вирусы (табл. 21-3). Они могут оказывать либо непрямое промоторное действие, либо способствовать неопластической трансформации инфицированных клеток. [c.466]
В отличие от ДНК-содержащих вирусов большинство ретровирусов (см. разд. 5.5.8 и 13.4.2) относительно безвредны для клетки-хозяипа. Зараженная клетка постоянно выделяет новые вирусные частицы. которые отпочковываются от плазматической мембраны. не вызывая неопластической трансформации клетки Однако изредка может происходить случайное овладение. захват ретровирусом регуляторного клеточного гена (или его испорченной копии, или фрагмента этого гена), который не используется в жизненном цикле самого вируса, но может кардинально влиять на судьбу клетки-хозяипа. В частности, как мы видели в гл. 13 (разд. 13.4.2), ретровирус, подобный вирусу саркомы Рауса. захватившему клеточный онкоген (рис. 21-21), легко обнаруживается по своему доминантному трансформирующему эффекту на инфицированные клет- [c.468]